Грибы – это одна из самых многочисленных групп организмов на Земле, которые играют важную роль в природных экосистемах и имеют большое значение для человека. Однако, грибы не все одинаковы, и существуют различные виды и разновидности, среди которых можно выделить несовершенные грибы.
Несовершенные грибы, или иных словах дефектные грибы, это грибы, не обладающие половым размножением. В отличие от совершенных грибов, которые производят споры, несовершенные грибы размножаются асексуально путем образования клеток-конидий или спорангиев. Они образуют грибницу, но не образуют споры, которые бы могли привести к половому размножению.
Несовершенные грибы встречаются в основном в гниющей древесине, почве, на растениях и в других местах. Они могут быть как паразитами, так и сапрофитами, то есть питаться за счет разложения органического материала. Некоторые несовершенные грибы являются патогенами, вызывающими заболевания растений, животных и даже человека.
Важно отметить, что несовершенные грибы не являются отдельной таксономической группой, они включаются в роды и семейства, общие для грибов в целом. Вместе с тем, многие несовершенные грибы являются источником ценных биологически активных веществ, которые находят применение в фармацевтической и пищевой промышленности.
Несовершенные грибы: что это, кто они и их разновидности [Грибы category]
Несовершенные грибы, или анаспории, представляют собой группу грибов, у которых отсутствует возможность образования сексуальных спор. Они получили свое название из-за этой особенности, их споры не образуются половым путем.
Кто такие несовершенные грибы?
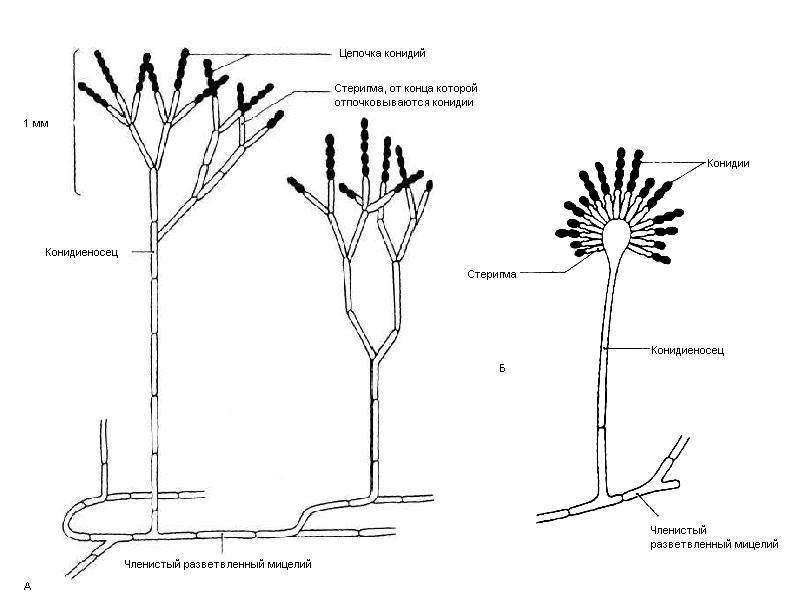
Несовершенные грибы образуют большую группу грибов, которые не входят в классическую систематику грибов с половым размножением. Вместо этого, они зависят от асексуального размножения, осуществляемого с помощью конидиоспор или дугоспор. Конидии могут быть свободными или содержаться в мешках на конидионосцах.
Однако несовершенные грибы являются важными растительными патогенами и вызывают различные заболевания в сельском хозяйстве. Они также могут быть полезными микроорганизмами, используемыми в промышленности для производства пищевых продуктов и лекарственных препаратов.
Разновидности несовершенных грибов
Существует множество разновидностей несовершенных грибов, включая такие роды как Aspergillus, Penicillium, Fusarium и Trichoderma. Они различаются по своим морфологическим и биохимическим характеристикам, а также по способу жизни и взаимодействию с окружающей средой.
Каждый род несовершенных грибов имеет свои уникальные свойства и может использоваться в различных отраслях. Например, некоторые роды несовершенных грибов могут производить антибиотики, а другие могут быть использованы для биологического борьбы с вредителями растений.
Понятие несовершенных грибов

К несовершенным грибам относятся многие виды патогенных или сапрофитных грибов, которые могут вызывать различные болезни у растений, животных и человека. Некоторые из них могут быть опасными для здоровья и могут вызывать аллергические реакции или инфекции.
Разновидности несовершенных грибов:

1. Аскомицеты (Ascomycetes) – грибы, которые образуют специальные клетки, называемые асками, внутри которых образуются аскоспоры.
2. Базидиомицеты (Basidiomycetes) – грибы, которые образуют базидии, внутри которых образуются базидиоспоры.
3. Циотериомицеты (Zygomycetes) – грибы, которые образуют зигоспоры, соединяющиеся вместе и образующие клетку-ресничку.
4. Дейтеромицеты (Deuteromycetes) – грибы, у которых не известны стадии образования спор, их размножение происходит только путем изображения и мицелиями.
Все эти разновидности несовершенных грибов имеют свои особенности и роли в биологических процессах. Понимание и изучение несовершенных грибов важно для понимания и поиска путей борьбы с патогенными грибами и развития применения полезных свойств грибов в медицине, пищевой промышленности и других областях.
Общая информация о несовершенных грибах
Несовершенные грибы представляют собой грибы, у которых отсутствует половой процесс или он происходит в очень ограниченных условиях. Такие грибы классифицируются в отдельную группу, отличную от совершенных грибов, у которых половой процесс происходит в полной мере.
Основной особенностью несовершенных грибов является их способность к размножению с помощью спор. Это позволяет им колонизировать и обживать различные среды и условия. Как и совершенные грибы, несовершенные могут образовывать грибницы, состоящие из мицелия – нитевидной структуры, способной распространяться и обеспечивать выживание грибов.
Разнообразие несовершенных грибов

Существует большое разнообразие несовершенных грибов, включая такие разновидности, как плесневые грибы (например, Aspergillus, Penicillium), дрожжи (например, Saccharomyces cerevisiae), а также другие болезнетворные грибы, вызывающие различные инфекционные заболевания у растений, животных и человека.
Важно отметить, что несовершенные грибы играют значительную роль в биотехнологии и медицине, используясь для производства различных продуктов, а также в процессе исследований и разработки новых препаратов.
Различные виды несовершенных грибов

Существует множество разновидностей несовершенных грибов. Некоторые из них являются паразитами растений и животных, например, рода Penicillium и Aspergillus. Эти грибы могут вызывать болезни у растений и животных, что приводит к снижению урожайности и заболеваниям.
Другие несовершенные грибы, например, роды Trichoderma и Gliocladium, являются противогрибковыми агентами. Они используются в сельском хозяйстве и садоводстве для борьбы с патогенными грибами. Несовершенные грибы также могут применяться в пищевой промышленности, например, роды Aspergillus и Penicillium используются для производства пищевых добавок и ферментов.
Несовершенные грибы представляют большой интерес для ученых и исследователей, так как они являются важными компонентами экосистем и имеют большое значение для сельского хозяйства, медицины и пищевой промышленности.
Уникальные свойства несовершенных грибов
У несовершенных грибов есть несколько уникальных свойств:
1. Отсутствие полного жизненного цикла: Одним из главных отличий несовершенных грибов от совершенных является наличие только анаспор с патогенностью. Это означает, что они не образуют фруктовых тел и спор или органов размножения, как это делают совершенные грибы.
2. Быстрое размножение: Несовершенные грибы обладают высокой способностью к размножению и быстрому распространению. Их споры быстро созревают и отсылаются на большие расстояния в окружающую среду.
3. Использование в биотехнологии: Несовершенные грибы часто используются в биотехнологических процессах для производства различных продуктов, таких как ферменты, витамины и антибиотики. Их способность к быстрому размножению и высокая активность ферментов делают их ценным ресурсом в промышленности.
4. Влияние на окружающую среду: Несовершенные грибы могут быть как полезными, так и вредными для окружающей среды. Некоторые несовершенные грибы являются патогенами для растений и животных, вызывая различные болезни. Другие же могут быть биоразлагателями, способными разлагать органические вещества и очищать окружающую среду от загрязнений.
5. Разнообразие видов: В настоящее время известно более 25 000 видов несовершенных грибов. Они представлены различными формами и размерами, от микроскопических грибов до крупных колоний на поверхности почвы.
В целом, несовершенные грибы являются важной группой организмов, которые играют значительную роль в биологическом образе жизни. Изучение их уникальных свойств помогает расширить наши знания о мире грибов и их роли в природе и на человеческое здоровье.
Распространение и местообитание несовершенных грибов
Несовершенные грибы широко распространены по всему миру и можно встретить их практически на всех континентах и во всех климатических зонах. Они обитают в различных местах и предпочитают разные условия для своего роста.
Местообитание несовершенных грибов
Несовершенные грибы могут быть найдены в самых разнообразных местах. Они могут обитать на почве, деревьях, плодах, органических отходах, растениях, животных и водных экосистемах. Их наличие и распространение в основном зависит от наличия подходящих условий для их размножения и роста.
Распространение несовершенных грибов
Несовершенные грибы могут распространяться различными способами. Одним из наиболее распространенных способов распространения является ветровая дисперсия. При этом споры несовершенных грибов могут передвигаться на большие расстояния с помощью ветра и оседать на различные поверхности, где они могут прорастать и размножаться.
Активное распространение несовершенных грибов также может осуществляться через животных. Животные, такие как насекомые, птицы и млекопитающие, могут переносить споры грибов на своих телах или внутри своих организмов и распространять их во время своей ежедневной деятельности.
Кроме того, несовершенные грибы могут распространяться через контакт с человеком. Человек может быть источником распространения спор грибов, их передачи на различные предметы, растения и другие организмы. Это может происходить при работе с почвой, растительной массой, зараженными растениями и другими биологическими материалами.
В общем, несовершенные грибы способны адаптироваться к разным условиям и распространяться в различных экосистемах, что делает их одними из наиболее разнообразных и успешных организмов на Земле.
Вопрос-ответ:
Что такое несовершенные грибы?
Несовершенные грибы – это грибы, которые не образуют половых спор. У них отсутствует стадия совершенного гриба, когда он производит споры.
Какие грибы относятся к несовершенным?
К несовершенным грибам относятся различные виды плесневых грибов, мукоровые грибы, дрожжеподобные грибы и др. Это широкая группа грибов, характеризующихся отсутствием полового размножения.
Какие разновидности несовершенных грибов существуют?
Существует множество разновидностей несовершенных грибов. Например, к разновидностям несовершенных грибов относятся Penicillium, Aspergillus, Fusarium и другие. Каждая разновидность имеет свои особенности и применяется в различных областях.
Какие особенности отличают несовершенные грибы от совершенных?
Главное отличие несовершенных грибов от совершенных заключается в отсутствии стадии полового размножения. Также, несовершенные грибы могут быть более устойчивыми к неблагоприятным условиям среды и иметь более широкий спектр питательных сред. Однако, несовершенные грибы не образуют спор, что делает их менее распространенными в природе.
Видео:
"Группа Несовершенные грибы", Биология 7 класс, Сивоглазов
ВСЁ, что нужно знать про ГРИБЫ для ЕГЭ по биологии
Знакомство с грибами Коровник Свинушка тонкая
Отзывы
Maria92
Статья очень интересная и информативная! Я, как любительница природы и грибов, с большим удовольствием узнала о несовершенных грибах. Оказывается, это группа грибов, которые отличаются от остальных своей неполноценной способностью к размножению. Интересно, что они не образуют созревших споров, как это делают другие грибы. Статья детально описывает различные виды несовершенных грибов, и это впечатляет! Кто мог подумать, что они настолько разнообразны? Есть грибы, которые образуют только одну клетку на сортах хлеба, а есть те, которые могут вызывать опухоли на растениях. Их разведение в лабораторных условиях представляется мне настоящим вызовом, но, наверное, очень увлекательным! Я благодарна авторам статьи за полезную информацию и за то, что они показали мне, что мир грибов на самом деле настолько разнообразен и удивителен. Теперь я буду обращать внимание на несовершенные грибы при своих лесных походах и интересоваться их различными разновидностями.
Niko61
Ах, эти несовершенные грибы! Когда я впервые услышал об их существовании, я был поражен. Кто бы мог подумать, что существуют грибы, которые не имеют полового размножения? Но так оно и есть. Несовершенные грибы – это разновидность грибов, которые не образуют споры, как их совершенные собратья. Это интересное явление в мире грибов, которое до сих пор вызывает много вопросов у ученых. Существует несколько разновидностей несовершенных грибов. Одним из наиболее известных и распространенных является аскомицет. Этот гриб имеет споры, но он не может размножаться половым путем, как большинство других грибов. Также есть разновидность грибов, которые имеют удлиненный цилиндрический вид и называются клависептины. Эти грибы также не могут размножаться половым путем. Интересно, что несмотря на то, что несовершенные грибы не могут размножаться половым путем, они все же являются известными паразитами многих растений. Они могут вызывать различные заболевания у растений и уменьшать их урожайность. Несовершенные грибы – это действительно удивительное явление природы. Они еще далеки от полного понимания учеными, и их изучение продолжается. Лично я увлекаюсь грибами, и эта новая информация о несовершенных грибах только увеличивает мое любопытство и желание узнать больше о них.
Alina_24
Эта статья оказалась мне очень полезной и интересной, так как я очень люблю готовить и экспериментировать с разными ингредиентами, в том числе и грибами. Всегда знала, что грибы являются ценным продуктом, но не знала, что они могут быть еще и “несовершенными”. Интересно узнать, что грибы с такими дефектами растут под влиянием вредных факторов, таких как грибные болезни, насекомые или погодные условия. Было интересно узнать о разновидностях несовершенных грибов, таких как “гнилые”, “искусственные” или “валяющиеся”. После прочтения статьи, я решила, что несовершенные грибы также могут быть полезными для приготовления разных блюд. Например, гнилые грибы можно использовать для приготовления супов или соусов, а искусственные грибы станут отличным ингредиентом для пиццы или пасты. Также, статья дала мне информацию о том, как правильно хранить грибы, чтобы избежать их порчи. Было полезно узнать, что грибы нужно хранить в прохладном, сухом месте, а также они быстро портятся, поэтому их лучше использовать как можно скорее после покупки. В целом, статья очень полезна для любителей готовить и экспериментировать с разными продуктами. Она дает информацию о несовершенных грибах и их разновидностях, а также рекомендации по их использованию и хранению. Теперь я точно знаю, что делать, если в магазине увижу несовершенные грибы!
Анна Иванова
Очень интересная статья! Я всегда была увлечена природой и часто отправляюсь в лес искать различные грибы. Но недавно я впервые услышала о таком явлении, как несовершенные грибы. Оказывается, это особый вид грибов, у которых происходит необычное смешение половых клеток. Мне было очень интересно узнать о различных разновидностях несовершенных грибов, таких как аскомицеты, блестиальные грибы и спороносные грибы. Я даже узнала, что некоторые из них встречаются преимущественно на определенных растениях. Теперь я с нетерпением жду следующей поездки в лес, чтобы поискать такие интересные и необычные грибы. Спасибо за информацию!